LA BECCACCIA SCIENTIFICA (a cura di Enrico Cavina)
ELEMENTI ANATOMO-FISIOLOGICI(correlati alla migrazione)
Il cervello della beccaccia è unico tra gli uccelli (Fergus Chuck-Wildlifenotes-Pennsylvania Game Commission-Harrisburg,2004-www.pgc.state.pa.us ).
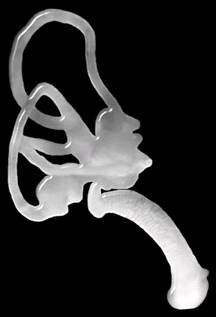
Il cervelletto – che controlla il coordinamento dei muscoli ed il bilanciamento del corpo – è collocato sotto il resto del cervello e sopra la colonna spinale. Una teoria ipotizza che nella evoluzione ancestrale corporea della beccaccia, gli occhi si sono via via collocati indietro nel cranio, il becco si è allungato e le narici e le orecchie si sono avvicinate alla base del becco, il tutto per consentire un miglior sondaggio del terreno dove trovare i vermi. Come risultato di questa dinamica evolutiva, il cervello è stato forzato all’indietro, il cervello medio e posteriore è stato spinto sotto e leggermente in avanti, così si può dire che oggi la beccaccia ha un “cervello capovolto”. Già in precedenza Stanley Cobb ( “On the angle of the cerebral axis in the American Woodcock” – Auk,vol.75,55-59,2959)aveva descritto i particolari di questa singolare conformazione del cervello e cervelletto, ed aveva evidenziato lo stretto rapporto tra bulbo olfattivo ed orbita, argomentando anche l’aspetto evolutivo del rapporto tra “grandi” occhi e becco: gli occhi sarebbero così grandi per facilitare le attività alimentari ed i movimenti notturni (controllo a 360° dei predatori) consentendo di vedere avanti e dietro, ed in alto per l’involo “a colonna” nel bosco, e sarebbero più riparati dal fango smosso dal becco.
Occhi, orecchie (e relativo orecchio interno con apparato vestibolare) ed apparato olfattivo hanno capacità di coordinarsi per le attività alimentari e più compiutamente per tutte le attività migratorie (orientamento, assetto di volo, scelte di direzione geografica e di sosta ecc.). Nel suo insieme questo coordinamento (riflessi ed elaborazioni dei circuiti nervosi ed endocrini) costituisce un eccezionale sistema sensitivo molto sviluppato nella beccaccia. Come già detto (Chuck F.-2004) “il cervello della beccaccia è unico tra gli uccelli “.
VEDI IMMAGINI TRATTE PER DIMOSTRAZIONE DIDATTICA DA:
people.eku.edu/ritchisong/birdbrain2.html
BIO 554/754 Ornithology
Nervous System: Brain and Special Senses II
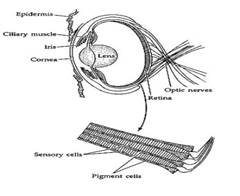
Struttura dell'occhio aviario:
- forma caratteristica, con sezioni anteriori e posteriori che sono sfere di raggi ineguali. Le forme degli occhi variano tra gli uccelli, che vanno da globose o appiattite (la maggior parte degli uccelli diurni; in basso) a tubolari (gufi; anche mostrato di seguito)>
- la luce che entra nell'occhio viene rifratta dalla cornea e dalla lente e colpisce la retina dove si trovano le cellule sensoriali (le cellule che convertono la luce in impulsi nervosi)
Le orecchie sono anch’esse molto sviluppate (10mm) e sono situate sul prolungamento craniale della parte inferiore del becco e sono sprovviste di padiglione auricolare. L’udito è acutissimo, verosimilmente capace di percepire ed interpretare anche gli ultrasuoni , ed è certamente specializzato nel percepire i rumori prodotti dai vermi nel terreno. Non si esclude che possa anche servire nel percepire segnali tra le beccacce (pericoli) ed anche segnali di gruppo in migrazione notturna.Per facilitare la perforazione e progressione “mirate” nel terreno, nel becco sono stati ben identificati organi tattili (corpuscoli di Herbst) costituiti da cellule sensoriali che percepiscono vibrazioni e modificazioni di pressione e quindi permettono di localizzare al meglio i vermi. Per la parte “meccanica” il becco ha intorno alla metà un sistema articolare di micro-ossicini capaci di fare da puleggia per poter aprire e divaricare nel terreno la parte terminale anterior-superiore del becco in modo da poter ingurgitare i vermi.L’orecchio interno con i canali semicircolari, saccula, otricolo, coclea gestisce il bilanciamento e l’equilibrio anche durante il volo , ed in particolare la coclea ,con il suo epitelio di cellule cigliate sensibile ai mutamenti di pressione e di livello del liquido in essa contenuto, ttiva i riflessi cerebellari che coordinano poi i movimenti muscolari utili alle necessità di volo . Queste generalità comuni agli uccelli e con caratteristiche di specializzazione più pronunciate per i migratori, lasciano intendere l’importanza dell’apparato cocleare attivo nel mantenimento dell’equilibrio direzionale ed altimetrico nei voli migratori della beccaccia e nelle capacità di volo nel bosco .
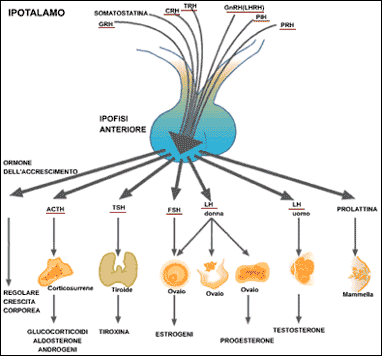
Nella scatola cranica strettamente connesse al cervello vi sono –come nell’uomo – due ghiandole endocrine ( Ipofisi ed Epifisi ) che producono ormoni : le loro attività,peraltro molto interconnesse tra loro ed il SNC, sono strettamente legate alla luce del giorno. Queste ghiandole sono al vertice di tutto il sistema endocrino ed esercitano un’azione determinante sulle attività di tiroide (TSH),surreni (ACTH),ghiandole sessuali (testicoli ed ovaie / LH,FSH);agiscono anche sul metabolismo esso stesso differenziato secondo l’età ed i cicli stagionali. Stimolano l’accrescimento ed attività delle ghiandole sessuali quando (luce del giorno che si allunga) gli uccelli nei quartieri invernali devono decidere di tornare ai quartieri di riproduzione . Sono invece inibite (luce del giorno che si accorcia) ed inibiscono a loro volta le funzioni sessuali quando gli uccelli devono prepararsi al volo autunnale verso i quartieri di svernamento: la tiroide, ghiandola in parte da loro dipendente, promuove l’accumulo di grasso ed il trofismo muscolare necessari per il lungo volo migratorio.L’Ipofisi è strettamente connessa con l’ipotalamo (SNC) ed è divisa in due parti (neuroipofisi e adenoipofisi), risente della luce che quindi è in grado di regolare la produzione di gonadotropine, gli ormoni che regolano il ciclo sessuale; è quindi evidente la connessione con gli stimoli migratori.
L’Epifisi o ghiandola Pineale, appartiene all’epitalamo ed è collegata con un intreccio di fasci nervosi alle circostanti parti nervose.
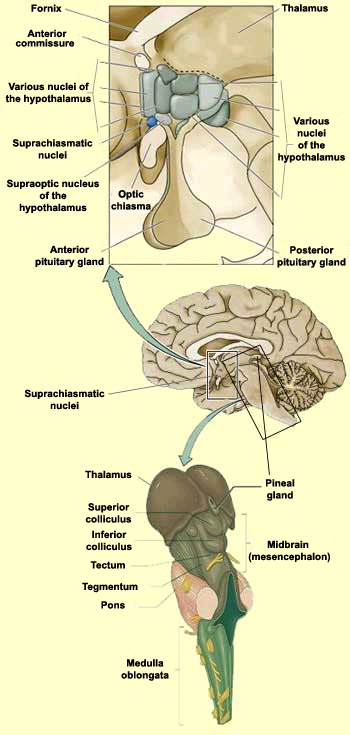
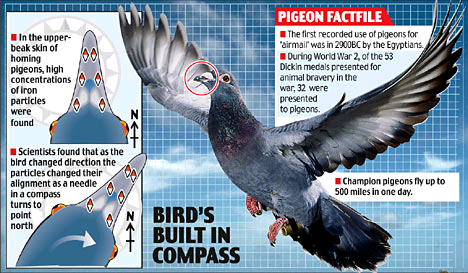
La ghiandola acquisisce dall’esterno e contiene anche un materiale magnetico (magnetite – ossido di ferro) e questo materiale in microcristalli endotissutali è considerato alla base delle capacità di navigazione migratoria degli uccelli (“bussola”) essendo capace di monitorare i campi magnetici dell’ambiente attraversato e quindi influire massivamente all’allineamento del corpo nello spazio. E’ stato accertato sperimentalmente che se si manipola la direzione e l’intensità dei campi magnetici intorno al capo degli uccelli, viene alterata la capacità di orientamento; inoltre viene inibita l’attività propria della ghiandola pineale e si riduce la produzione di melatonina; da ciò deriva lo sconvolgimento dei ritmi circadiani.
La ghiandola, ritenuta anche fotosensitiva di per se e per i riflessi con la retina, ha un ruolo essenziale nel determinismo degli stimoli migratori; questi stimoli si coordinerebbero con i materiali magnetici contenuti nella ghiandola e questi si allineerebbero ai campi magnetici terrestri realizzando come un cartello indicatore (“road sign” ) per l’orientamento migratorio (Csernus V.,Mess B. – Biorhytms and pineal gland – Neuroendocrinology Letters 6,24,404-411,2003 – www.nel.edu). Altri recentissimi studi hanno evidenziato capacità di adattamento e controllo delle attività della retina sino a poter regolare cambiamenti di stile di vita “notturna” durante la migrazione, ne riportiamo un elenco:
- Shocklet – Comparison of circadian changes in the retinas of migrating and non-migrating birds – Thesis-Undergraduate Research Texas A-M University – April 2006 – www.TexasA-MUniversity
- -Wiltschko&R. – Magnetic compass orientation in birds and its physiological basis –Naturwissenchaften 89,445-452,2003
- Kusnar e al. – The bird clock :a complex multi-oscillatory and highlydiversified system.- Bio.Rhytm Res.35,121-144,2004
- MouritsonU. e al.- Cryctochromes and neuronal activity markers colocalize in the retina of migratory birds during magnetic orientation – Proc.Nat.Acad.Sci.USA 101,39,14294-14299,2004 .
Ancora altri recenti studi (Fusani L.<Univ.Siena> & Gwinner E. – Melatonin and Nocturnal Migration – Annali NY Avad,Sci. 1046,264-270,2005 – www.annalsnyas.org) evidenziano che i livelli notturni di melatonina prodotti dalla ghiandola Pineale sono più bassi durante tutto il periodo migratorio (dormono di meno durante la notte) e direttamente od indirettamente contribuiscono a regolare le attività notturne per la reintroduzione delle riserve alimentari energetiche necessarie in misura diversa nei periodi migratori ed in quelli di permanenza di riposo. Certamente queste implicazioni riguardano anche le abitudini ed attività notturne della beccaccia durante la migrazione e poi lo svernamento. Studi specifici (Ferrand) utilizzano un Indice di Attività Notturna (I.A.N.) come vedremo in seguito (ndA).Queste citate ricerche indagano – in diverse condizioni di stimoli e relativi riflessi – sulle condizioni funzionali delle strutture anatomiche deputate quali centri nervosi,nervi,epiteli sensitivi situati nelle ghiandole endocrine endocraniche e nella retina. Da tutti questi lavori, per lo più recenti nell’ultimo decennio, si evince come sia attuale la ricerca nel settore “migrazione degli uccelli” anche per gli aspetti morfo-funzionali .Chiunque si accinge ad esplorare quello che ancora è definito il “mistero” delle migrazioni dovrebbe – a nostro avviso- applicarsi ad uno sforzo conoscitivo il più possibile aggiornato ed approfondito.Come già accennato in precedenza, gli elementi della retina (alcuni elementi più specificatamente dedicati) hanno funzione di recettori che si attivano con fotopigmenti (oggi sempre più identificati come criptocromi) insieme ad altro tipo di recettori che sono stimolati dalla magnetite (minerale ferroso- Ossido di Ferro –“Fe3O4 “– ioni metallici tipo A e B in cristalli). Questi recettori sono strettamente legati con la branca oftalmica del nervo trigemino e sono sensibilissimi anche a minimali cambiamenti del campo magnetico.
Come queste azioni di stimoli e riflessi si coordinano tra loro, rimane ancora sconosciuto. (Beason R.C. – Mechanism of magnetic orientation in birds – Integr.Comp.Biology 45,3,565-573,2005 – www.bione.org ).ndA: questi elementi relativi alla percezione del campo magnetico sono certamente da tenere in considerazione per quanto riguarda l’inquinamento magnetico (magnetic pollution) da ripetitori di telefoni cellulari, antenne varie TV e Radar e relativi fenomeni di disorientamento degli uccelli migratori (vedi in seguito).Negli uccelli la magnetite è stata isolata nei tessuti del capo, particolarmente nella regione etmoide situata alla base del becco e nella cute della mandibola superiore, tutte zone che sono innervate dal nervo oftalmico (trigemino).
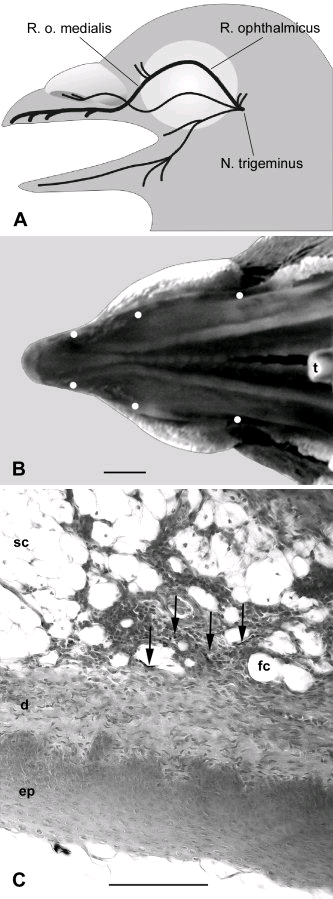
A questi reperti si aggiunge l’identificazione ultrastrutturale di cellule che contengono magnetite. I micro-cristalli di magnetite possono risentire, in una specie di catena assiale, delle interazioni dal campo magnetico, ruotando ed allineandosi in una forma di movimento di compasso. Alcuni esperimenti realizzati modificando il campo magnetico dimostrerebbero che negli uccelli migratori questi magneto-recettori si orienterebbero secondo specifici assi direzionali, ad esempio est-ovest. ( Wiltshko W.e al. – Magnetite-based magnetoreception in birds : the effect of biasing field and a pulse on migratory behavior – Journ.Experim.Biol. 205,3031-3037,2002 ). Studi più recenti (2005) dimostrerebbero che esistono due diversi sistemi sensibili al campo magnetico:
- uno basato sulla magnetite presente nei tessuti alla base del becco (vedi qui figura – piccione)
- uno basato su processi di elaborazione dipendenti dalla luce.In tal senso questi rilievi dimostrebbero
(Mouritsen H.& Ritz T. – Magnetoreception and its use in bird’s navigation – Current Opinion Neurobiology 15,4,406-414,2005) - le risposte comportamentali degli uccelli dipendenti dall’oscillazione del campo magnetico;
- la presenza di sensori putativi (criptocromi) negli occhi dei migratori;
- un’area del cervello specializzata ad integrare i suddetti fattori con gli “input” visivi notturni nei migratori notturni;
- un’altra area putativa, sensibile al magnetismo, situata alla base del becco.
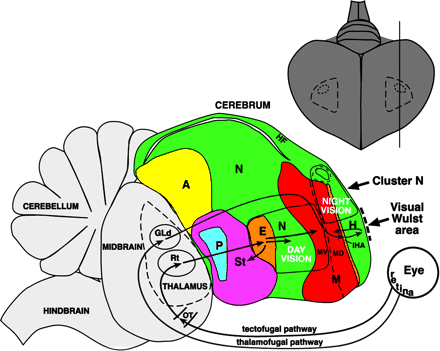
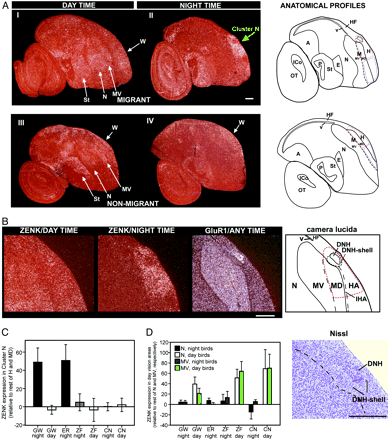
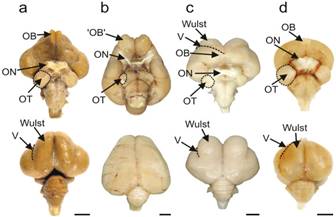
Altre immagini didattiche delle aree visive di uccelli da (Graham R. Martin e al.-Kiwi Forego Vision in the Guidance of Their Nocturnal Activities - PLoS ONE. 2007; 2(2): e198.)
Visual processing areas of the brains of four species of birds. Ventral and dorsal views of the brains of
a: Emu (diurnal, flightless);
b: Kiwi (nocturnal, flightless);
c: Barbagianni (nocturnal, flying),
d: Colombo (diurnal, flying).
OT: optic tectum; ON: optic nerve ; OB: olfactory bulb (che in realtà consiste in un foglio corticale simile nel kiwi adulto) – V: vallecula.
Nota il diametro ridotto del nervo ottico nel Kiwi rispetto a quello delle altre tre specie (vedi testo per le misurazioni effettive). Nella vista dorsale del Kiwi, nota l'estensione caudale dei grandi emisferi telencefalici, che nascondono completamente il mesencefalo sottostante. Si noti anche nel Kiwi che non esiste un evidente rigonfiamento sul dorso dell'emisfero che identifica il Wulst in specie come Barn Owl ed Emu. Barre di scala: Emu, 1 cm; Kiwi, Barbagianni e Colombo: 0,5 c
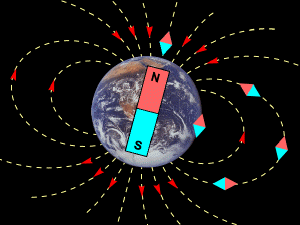
Beason R.C. nel basilare lavoro del 2005 (Beason R.C. – Mechanism of magnetic orientation in birds – Integr.Comp.Biology 45,3,565-573,2005 – www.bione.org ) sottolinea come l’orientamento migratorio visivo può entrare in crisi in condizioni di cielo coperto,nebbia,nubi basse. Il campo magnetico terrestre però è sempre presente in ogni luogo del pianeta e “penetra” in ogni struttura liquida, gassosa, solida incluso il corpo degli animali. Queste caratteristiche fanno sì che il campo magnetico può sempre fornire,ovunque,informazioni direzionali. Gli uccelli usano il campo magnetico come un compasso, ma sono anche sensibili a possibili variazioni temporali e spaziali.
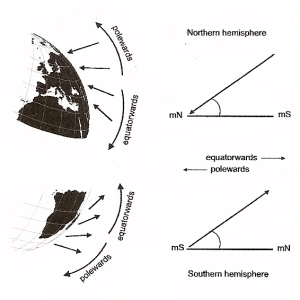
L'inclinazione della bussola aviaria fornisce informazioni sull'allineamento del campo magnetico e non sulla polarità del campo (come fa una bussola). Insieme ai rilievi visivi gli uccelli migratori sono anche capaci di usufruire di una “mappa” magnetica dove possono rilevare variazioni - anche molto piccole - del campo magnetico che stanno sorvolando, ed anche in dipendenza del livello di latitudine ( in discesa a Sud , in risalita a Nord ) appunto più o meno vicino all’equatore.
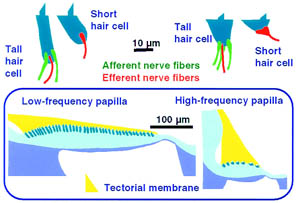
Anche l’apparato vestibolare (orecchio interno) deputato all’equilibrio direzionale risente di queste variazioni.La coclea è il punto in cui il "suono" viene convertito in impulsi nervosi che viaggiano nel cervello. Più precisamente, le cellule ciliate (papilla basilare) all'interno della coclea sono "deviate" dalle onde di pressione, generando impulsi nervosi
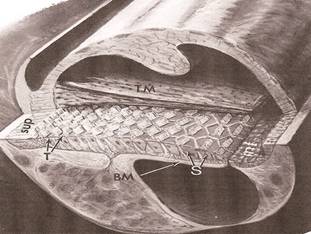
Coclea aviaria. L'epitelio sensoriale, costituito da cellule ciliate e cellule di supporto, poggia sull'elemento basilare (BM). La cellula dei peli varia in altezza da alta (T; vicino al margine superiore, sup) a corta (S; vicino al margine inferiore, inf). La membrana tettorale (TM) si trova appena sopra i peli (chiamata anche stereocilia). Le aree "vuote" al di sopra della membrana tettorale e al di sotto della membrana basilare sono riempite con un fluido chiamato perilinfa (Figure from Tilney and Saunders 1983).
Gli uccelli godono di una visione più nitida rispetto agli umani. Gli uccelli possono vedere determinate frequenze di luce - incluso l'ultravioletto - che gli umani non possono vedere. In effetti, molti uccelli canori hanno piume che riflettono la luce ultravioletta. Questa luce viene utilizzata per comunicare specie, genere e forse anche posizione sociale. Gli uccelli possono vedere questa luce ultravioletta in normali condizioni di luce diurna. Gli umani richiedono l'assistenza di una luce nera. Perché gli uccelli vedono meglio degli umani?

1) Sia gli uccelli che gli umani hanno "coni" fotorecettivi nella retina situata nella parte posteriore dell'occhio. Questi coni ci permettono di vedere la luce colorata. L'occhio umano contiene 10.000 coni per millimetro quadrato. Gli uccelli selvatici hanno fino a 12 volte questa quantità o 120.000 coni per millimetro quadrato.
2) Nell'uomo, questi coni fotorecettivi sono costituiti da tre tipi. Ogni cono è sensibile alla luce rossa, verde o blu. Questo si chiama visione dei colori tricromatica. Gli uccelli hanno un cono in più per la visione quadricomatica dei colori. Questo cono extra espande lo spettro della luce visibile, permettendo agli uccelli di vedere le frequenze ultraviolette.
3) In condizioni di scarsa luminosità, sia gli umani che gli uccelli fanno affidamento su "bastoncini cellulari" fotorecettivi nella retina. L'occhio umano ha 200.000 barre cellulari per millimetro quadrato. Alcuni uccelli, come i gufi, hanno fino a 1.000.000 di aste cellulari per millimetro quadrato.
4) Gli occhi degli uccelli, in media, rappresentano il 15% della massa dell'intera testa dell'uccello. Gli occhi umani, al contrario, rappresentano meno del 2% della testa.
5) Le retine degli uccelli, contrariamente agli umani, non contengono vasi sanguigni. Questo impedisce la dispersione della luce e quindi fornisce agli uccelli una maggiore acuità visiva rispetto agli umani.
Volendo cercare di semplificare molto approssimativamente il problema “capacità direzionale” del fenomeno migratorio , si potrebbe dire che micro-lancette magnetiche ( micro-cristalli di magnetite) situate in strutture tissutali , si orientano in asse tra loro in dipendenza del campo magnetico terrestre , e trasmettono le informazioni di rotta che poi il cervello “vede” e decide di far seguire con il volo ( comandi ai muscoli) migratorio; il cervello usufruisce ovviamente di altre informazioni visive (sole,orizzonte,stelle,orografia) e comunque sensitive (olfatto,calore,udito) e centrali per gli adulti (memoria). Abbiamo più volte fatto riferimento alla retina ed alla sua struttura cellulare così importante anche per l’attivazione del compasso magnetico .La vista e gli occhi della beccaccia sono ben descritti nel vecchio testo di Califano ( 1° ediz.1971): “ gli occhi della beccaccia contrariamente a come molti credono non sono mobili, ma fissi nella loro cavità orbitale (sono le tre palpebre che si muovono –ndA),“con i due occhi riesce ad abbracciare un campo addirittura superiore a 360°”, “la parete posteriore dell’occhio è appiattita in modo che il campo visivo sia sempre a fuoco. Sulla Retina vi sono numerosi elementi visivi per unità di superficie e si presume che siano cellule specializzate nella visione dei colori (ndA: siamo nel 1971)“, “La pupilla ha proprietà veramente eccezionali dovendo l’uccello adattarsi ad ambienti scarsamente illuminati”. Molti lavori più recenti – anche testi di descrizione sistematica (Nervous System:Brain and Special senses II, BIO 554-75 Ornithology http://people.eku.edu/ritchsong/birdbrain2.html - 2006) - evidenziano l’eccezionale campo visivo della beccaccia a 360° e documentano la capacità degli elementi retinici per la discriminazione dei colori. La retina ha recettori di colori, recettori di bianco e nero ed un altro tipo di recettore capace di regolare i ritmi circadiani e questo gioca un suo ruolo nel processo di analisi dei segnali magnetici. (Ritz T. e al. – A model for photoreceptor-based magnetoreception in birds – Byophysical Journal 78,707-718,2000).
Volendo cercare di semplificare molto approssimativamente il problema “capacità direzionale” del fenomeno migratorio , si potrebbe dire che micro-lancette magnetiche ( micro-cristalli di magnetite) situate in strutture tissutali , si orientano in asse tra loro in dipendenza del campo magnetico terrestre , e trasmettono le informazioni di rotta che poi il cervello “vede” e decide di far seguire con il volo ( comandi ai muscoli) migratorio ; il cervello usufruisce ovviamente di altre informazioni visive ( sole,orizzonte,stelle,orografia) e comunque sensitive ( olfatto,calore,udito) e centrali per gli adulti (memoria). Abbiamo più volte fatto riferimento alla retina ed alla sua struttura cellulare così importante anche per l’attivazione del compasso magnetico .
La vista e gli occhi della beccaccia sono ben descritti nel vecchio testo di Califano ( 1° ediz.1971) : “ gli occhi della beccaccia contrariamente a come molti credono , non sono mobili , ma fissi nella loro cavità orbitale ( sono le tre palpebre che si muovono –ndA) , “ con i due occhi riesce ad abbracciare un campo addirittura superiore a 360°”, “ la parete posteriore dell’occhio è appiattita in modo che il campo visivo sia sempre a fuoco .Sulla Retina vi sono numerosi elementi visivi per unità di superficie e si presume che siano cellule specializzate nella visione dei colori (ndA:siamo nel 1971) “, “ La pupilla ha proprietà veramente eccezionali dovendo l’uccello adattarsi ad ambienti scarsamente illuminati”.
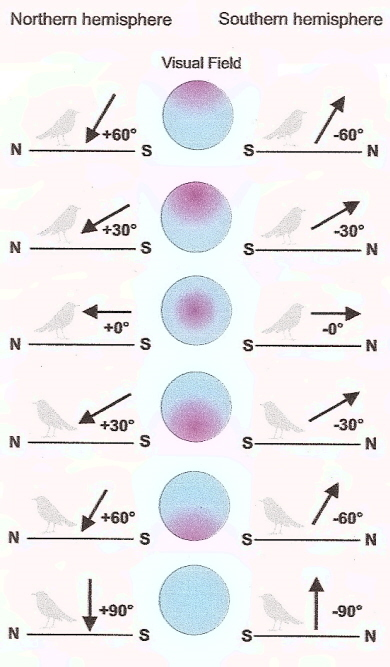
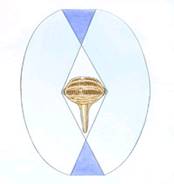
Bussola magnetica dipendente dalla luce
I campi magnetici esterni possono influenzare i processi indotti da fotoni che coinvolgono reazioni biomolecolari e recenti lavori suggeriscono che sono coinvolte molecole chiamate criptocromi. I fotorecettori magneto-sensibili distribuiti nella retina dell'occhio possono mostrare risposte aumentate o diminuite alla luce, a seconda del loro allineamento rispetto al campo magnetico e consentire agli uccelli di vedere effettivamente le linee del campo magnetico. Le prove attuali suggeriscono che l'occhio destro degli uccelli è più importante di quello sinistro nel "vedere" le linee del campo magnetico. La figura a sinistra mostra come le informazioni sulla magnetorecezione possano essere percepite dagli uccelli. L'uso di una bussola d'inclinazione può creare problemi vicino ai poli magnetici (90 gradi) e all'equatore magnetico (0 gradi). L'allineamento delle linee di campo (verticale ai poli e orizzontale all'equatore) rende impossibile scegliere la direzione corretta. Fortunatamente, gli uccelli hanno mezzi alternativi per ottenere informazioni direzionali (ad es. Sole e stelle). Da: Muheim (2004).
Usando il campo magnetico terrestre per navigare
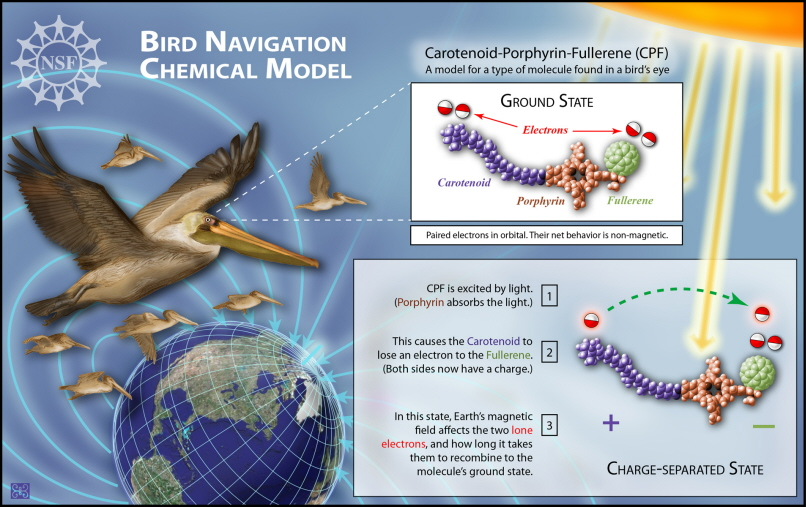
Le prove attuali suggeriscono che gli uccelli usano reazioni chimiche magneticamente sensibili avviate dalla luce (chiamata percezione magnetica chimica) per orientarsi, ma nessuna reazione chimica ha dimostrato di rispondere a campi magnetici deboli come quelli terrestri. Tuttavia, Maeda et al. (2008) hanno ora sintetizzato una molecola (carotenoide-porfirina-fullerene o CPF) che è sensibile sia alla grandezza che alla direzione dei minuscoli campi magnetici come quelli della Terra (che è, in media, un ventimillesimo forte quanto un magnete da frigorifero). La molecola sintetizzata è composta da tre unità (una triade carotene-porfirina-fullerene). Quando eccitato dalla luce, la molecola della triade forma uno stato separato dalla carica con la carica negativa sulla porzione di fullerene (o buckyball) simile a una palla da calcio e la carica positiva sulla porzione di carotene a forma di bastoncino. Si potrebbe pensare che la molecola della triade, nel suo stato separato di carica, abbia piccoli magneti a barra ad entrambe le estremità - così distanti che interagiscono tra loro solo debolmente.
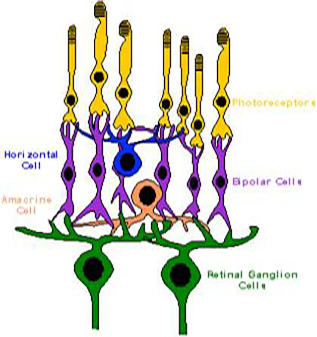
Molti lavori più recenti – anche testi di descrizione sistematica (Nervous System:Brain and Special senses II, BIO 554-75 Ornithology http://people.eku.edu/ritchsong/birdbrain2.html - 2006) - evidenziano l’eccezionale campo visivo della beccaccia a 360° e documentano la capacità degli elementi retinici per la discriminazione dei colori. La retina ha recettori di colori, recettori di bianco e nero ed un altro tipo di recettore capace di regolare i ritmi circadiani e questo gioca un suo ruolo nel processo di analisi dei segnali magnetici. (Ritz T. e al. – A model for photoreceptor-based magnetoreception in birds – Byophysical Journal 78,707-718,2000). La retina aviaria ha tre tipi di fotorecettori che "traducono" la luce in impulsi nervosi:• canne - visione in bianco e nero• coni - visione a colori• doppi coni - visione a colori Sono anche riconosciute cinque principali classi di neuroni retinici: fotorecettori, cellule bipolari, cellule orizzontali, cellule amacrine e cellule gangliari. I fotorecettori (bastoncelli e coni), cellule bipolari e cellule orizzontali stabiliscono contatti sinaptici con l'esterno strati retinici. Le cellule bipolari, amacrine e gangliari entrano in contatto con gli strati retinici interni.
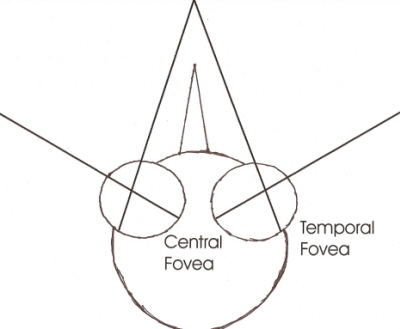
E’ molto significativa l’affermazione di Beason R.C. e che è generica per tutti gli uccelli “ Paragonati alle capacità visive degli uomini , gli uccelli possono distinguere più colori e recepire direttamente la luce ultravioletta e la luce polarizzata “. (Beason R.C. – Through a Bird’s Eye : exploring avia sensory perception . 2003 – Bird Strike Comm-Usa/Canada ). Ma viene da domandarsi : “la percezione della luce polarizzata e della luce ultravioletta è così importante per l’attivazione dei meccanismi direzionali negli uccelli migratori ? “La risposta viene in buona parte dalla sintesi di Mueller H. (Mueller H. – The sixth sense of Direction – Sun 12,17-19,2006 // - Navigation Secrets of Migratory Birds).
Tutte queste implicazioni/integrazioni – nello specifico della beccaccia – avrebbero anche grande importanza nel determinismo e controllo del volo migratorio notturno.Un lavoro molto importante e specifico sull’occhio della beccaccia è quello di Martin G.R. – Visual fields in woodcocks (Scolopax Rusticola) – Journal of Comparative Physiology 174,6,787-793,1994 –www.springerlink.com/ - dove si specificano importanti elementi conoscitivi.
- Sono assenti movimenti dell’occhio significativi;
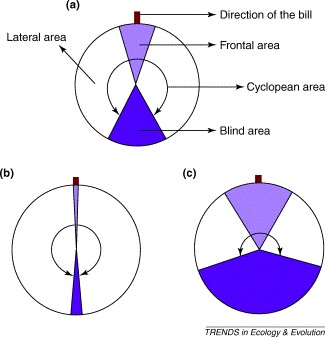
- Il campo retinico binoculare è lungo e ristretto; si estende per 190° nel piano sagittale Quando la testa si pone in una normale postura (becco in angolo di 40° rispetto al piano orizzontale) il campo binoculare si stringe da 25° sopra il becco a 5° sopra il piano orizzontale dietro la testa. Stando così le cose le beccacce hanno una copertura di visuale emisferica sopra di loro, ma il becco impedisce in basso la visione completa del terreno.
- I campi retinici monoculari coprono un angolo di 182° sul piano orizzontale (360° per la visione binoculare) e non vi sono settori ciechi dei campi ottici.
- La struttura del cranio della beccaccia facilita la visione panoramica sul piano orizzontale.
- Nel campo dell’anatomia comparata la configurazione del sistema visivo è più finalizzata alla visione utile per mangiare. La copertura visiva più vasta dell’emisfero celeste nella sua interezza si realizza solo in specie –come la beccaccia – che usano altri sensi (udito-olfatto) per guidare maggiormente l’atto alimentare ( Martin G.R.). (Hall M.I. e al.- Eye shape and activity pattern in birds – Journal pf Zoology 271,437-444,2007 –// - IwaniukN. e al.- The Comparative Morphology of the Cerebellum in Caprimulgiform Birds: Evolutionary and Functional Implications - Brain Behav Evol 2006;67:53-68 (DOI: 10.1159/000089120)